

Аннотация
Акклиматизировавшиеся инвазивные виды представляют собой постоянную угрозу биоразнообразию и продовольственной безопасности, и поэтому требуют выработки устойчивого подхода с целью смягчения отрицательных последствий. По общему мнению, широко распространившийся эктопаразитарный клещ Varroa destructor, инвазивный вид из Азии, стал основной биологической угрозой мировому пчеловодству на основе пчелы Apis mellifera. V. destructor почти полностью уничтожил популяцию дикой европейской медоносной пчелы (Apis mellifera). На сегодняшний день единственное средство в пчеловодстве – это постоянные меры контроля популяции клеща в течение сезона (противоклещевые обработки), что мешает процессу адаптации пчелы к паразиту.
Усилия по целенаправленной селекции до сих пор не привели к выведению толерантных или резистентных пчёл. Тем временем естественный отбор продемонстрировал свою успешность, по крайней мере, в семи разных, известных нам местах. В данной работе мы предлагаем воспользоваться преимуществами естественного отбора медоносных пчёл на устойчивость путём отказа от противоклещевых обработок на пасеках. Основные принципы данного подхода: спаривание собственных маток и трутней внутри популяции, а также отбор, основанный на критериях выживания и быстрого размножения пчелосемей.
Данный подход используется нами 10 лет, и его результатом стало появление «нормальных» пчелосемей с высоким уровнем резистентности к V. destructor. В этой работе мы призываем учёных и пчеловодов на местах присоединиться к новой программе естественного отбора, которая уже началась как минимум в трёх местах. Это в конечном итоге приведёт к появлению в мире нескольких популяций медоносных пчёл, хорошо адаптированных к местным условиям и с высокой резистентностью к V. destructor, и поможет стабилизировать глобальное пчеловодство.
Введение
Западная медоносная пчела (Apis mellifera L.) распространена почти по всему миру, как в дикой природе (является эндемиком в Европе, Африке и на Ближнем Востоке, натурализовалась в Америке, Азии и Австралии), так и на культурных пасеках (Moritz et al., 2005). A. mellifera является самым важным опылителем, которого использует человек, и таким образом вносит существенный вклад в глобальную продовольственную безопасность и производство продуктов пчеловодства (мёд, пыльца, воск и прополис, Klein et al., 2007; Aizen et al., 2008). Очевидно, что пчеловодство имеет большую экономическую и общественную значимость.
В результате глобализации пчеловодства увеличилось число новых патогенов, поражающих A. mellifera, переселившихся на медоносную пчелу с близкородственных видов. Так случилось с эктопаразитарным клещом Varroa destructor (Anderson and Trueman, 2000), который перешёл с восточной медоносной пчелы (Apis cerana) на A. mellifera.
В настоящее время этот клещ имеет почти повсеместное распространение (Ellis and Munn, 2005) и является основной биологической причиной гибели семей A. mellifera во всем мире (Neumann and Carreck, 2010; Rosenkranz et al., 2010). V. destructor является очень опасным переносчиком нескольких вирусов медоносных пчёл, вызывающих эпидемию внутри семьи, из-за чего семья приходит в упадок и погибает в течение 2-3 лет (Neumann et al. 2012).
Глобальное распространение V. destructor произошло очень быстро, и почти не оставило времени для смягчения последствий путём предотвращения интродукции паразита в новые регионы, уничтожения уже интродуцированных клещей, или, наконец, путём замедления его дальнейшего распространения (Van der Weijden et al., 2007).
Интересно, что A. mellifera – единственный вид медоносной пчелы, который изначально, до нашествия V. destructor, не имел клещей, паразитирующих на расплоде (Eickwort, 1994). Явное отсутствие истории совместной эволюции хозяина и паразита (Woolhouse et al., 2005) может объяснить гораздо более высокую восприимчивость A. mellifera к V. destructor и, как следствие, быстрое сокращение популяций диких колоний европейской медоносной пчелы и её гибридов во всём мире с начала распространения V. destructor (Rosenkranz et al., 2010; Villa et al., 2008; Kraus and Page, 1995).
Несмотря на то, что естественный отбор в случае прекращения лечения пчёл неизбежно привёл бы к адаптации A. mellifera к V. destructor, этот вариант в пчеловодстве не рассматривался, поскольку предполагаемая потеря пчёл вызывала понятную озабоченность по поводу устойчивости опыления сельскохозяйственных культур и, следовательно, продовольственной безопасности (Klein et al. 2007; Aizen et al. 2008). По этой причине V. destructor находится под постоянным контролем на пасеках, что предотвращает процесс коэволюции (как предсказывал Олдройд, Oldroyd, 1999), который ведёт к выработке резистентности и толерантности хозяина или к снижению вирулентности паразита (Neumann and Blacquie’re, 2017; Brosi et al., 2017).
Есть наглядные свидетельства того, что благодаря естественному отбору дикие популяции А. mellifera могут выжить после заражения V. destructor (Seeley, 2007; Fries et al., 2007; Fries et al. 2006; см. обзор у Locke, 2016). Это относится также к культурным пасекам, на которых пчёл не лечат от клеща (Oddie et al., 2017, 2018; Kruitwagen et al., 2017; Panziera et al., 2017; McMullan, 2018). Таким образом, устойчивое пчеловодство без необходимости лечения V. destructor бесспорно осуществимо. Даже несмотря на то, что механизмы, лежащие в основе устойчивости к V. destructor, до конца ещё не изучены, мы предлагаем использовать преимущества Дарвиновского естественного отбора для адаптации пчёл к этому новому паразиту как неотъемлемую часть местной пчеловодческой практики.
Мы предлагаем краткое описание метода естественного отбора, который можно вводить в практику пчеловодства любой местности — он включает в себя способы предотвращения возможного побочного ущерба, такого как, например, повторное заражение соседних пасек. Программа, получившая название «Пчелиный чёрный ящик Дарвина» (Darwin’s Black Bee Box’ – DBBB), была разработана на основе более ранних работ за 10 лет (Panziera et al., 2017; Kruitwagen et al., 2017). Эта программа будет проходить дальнейшее тестирование в новом проекте, который только что стартовал в трёх местах (т. е. с тремя локальными группами пчелосемей). Мы призываем пчеловодов и исследователей присоединиться к этому проекту и совместно следовать плану, изложенному ниже, в своих регионах.
«Пчелиный чёрный ящик Дарвина» (DBBB)
Предлагаемая программа основана на принципах эволюции посредством естественного отбора: пчелосемьи, способные выживать и успешно размножаться, передают свои гены следующему поколению. Аллели генов, кодирующих признаки, способствующие выживанию, будут постепенно встречаться всё чаще, в то время как присутствие вредных или нейтральных аллелей будет снижаться.
Как и Дарвин, мы можем наблюдать адаптацию, не понимая лежащих в её основе механизмов. Здесь мы используем аналогию с «чёрным ящиком», содержимое которого остаётся скрытым, тем не менее, результаты этого содержимого очевидны и видны.
Более того, данный подход заранее не выделяет те или иные качества пчёл, которые обычно считаются полезными (ХПП — хозяйственно полезные признаки), а просто следует за природой пчёл, стремящихся к выживанию и размножению. Внутри «чёрного ящика» аллели, связанные с жизнеспособным фенотипом, сохраняются и передаются следующему поколению. Таким образом, естественный отбор – это комплексный процесс, поскольку он поддерживает генетическое разнообразие, сохраняя все жизнеспособные фенотипы в «чёрном ящике», включая, возможные редкие аллели, полезные для выработки устойчивости к паразитам и патогенам (Delaplane et al., 2015).
Целенаправленная селекция пчёл, напротив, по определению снижает генетическое разнообразие, отбирая из выживших фенотипов только те, которые отвечают предпочтительным качествам, тем самым потенциально исключая многие фенотипы, несмотря на их способность к выживанию (Uzunov et al., 2017).
DBBB-программа учитывает естественный сезонный цикл размножения пчелиных семей, имитируя роение путём деления пчелосемей. Следовательно, популяция сохраняет не только своё генетическое разнообразие, но и сопутствующую разнообразную биоту (как полезные, так и вредные ассоциированные организмы: бактерии, дрожжи, грибы, вирусы, клещи и т.д.), что способствует уменьшению вирулентности паразита и восприимчивости хозяина (Neumann and Blacquie’re, 2017; Blacquie’re and Panziera, 2018).
Это также исключает ситуацию, когда матка следующего поколения попадает в совершенно незнакомую биотическую среду (новая семья + новая биота), как это бывает при подсадке покупных маток. В свою очередь расплоду семьи (и биоте) не приходится сталкиваться с чужой биотой, привнесённой молодой сторонней маткой.
Ниже мы описываем в структурированном виде практические принципы и методы нашего подхода, который мы активно пропагандируем и предлагаем использовать везде, где это возможно.
Условия, необходимые для реализации DBBB-программы
1. 25-30 пчелосемей
Программу DBBB можно начать весной на пасеке из 25-30 генетически разнообразных местных пчелосемей (от разных пчеловодов) из одного региона. Это обеспечит достаточную вариативность и локальную/региональную адаптацию в популяции (взаимодействие генотипа и окружающей среды, Büchler et al., 2014).
Успешно развивающиеся семьи нужно размножить на четыре отводка (см. план действий ниже), после чего в первый год эти семьи нужно однократно обработать против V. destructor щавелевой кислотой в период, когда молодые матки начали кладку и до запечатывания ячеек расплода. Семьи, которые переживут первую зиму и покажут хорошее развитие будущей весной (быстрый рост и вывод трутней), станут родительскими для производства следующего поколения.
В середине зимы, когда берут пробы пчёл для определения степени заклещёванности, четверть семей, размноженных отводками, можно отобрать в качестве контрольной группы. Наличие контрольной группы пчелосемей не обязательно, но может быть очень полезно в будущем для сравнения отобранных и не отобранных групп и выявления местных механизмов, обеспечивающих выживание семей в условиях заражённости V. destructor.
2. Изолированный район
Изолированность важна в начале лета для соблюдения условия спаривания маток и трутней внутри популяции. Если создаётся контрольная группа семей, то потребуется отдельное изолированное место. Достаточно изолированным можно считать место, если в радиусе 3 км от него нет или почти нет других пасек. Согласно исследованиям Яффе (Jaffe et al., 2009) и Морица (Moritz et al., 2007), при изолированности в радиусе 1.2 км 60-75% спариваний происходит с трутнями своей популяции. Полевые исследования Дженсена (Jensen et al., 2005) показали, что радиус спаривания (радиус полёта маток + радиус полёта трутней) в 50% случаев был меньше 2,5 км, а в 90% случаев спаривания происходили в пределах 7,5 км. Второе более крупное значение, по всей видимости, обусловлено тем, что матки увеличивают дальность вылетов, предположительно из-за нехватки трутней в данном районе, что в свою очередь снижает количество спариваний на каждую матку (Neumann et al., 1999а, b).
Предложенная нами DBBB-программа основана на отрицательном отборе (отход неадаптированных фенотипов) и не нацелена на усиление отдельных, выбранных человеком качеств и фенотипов. Поэтому, если несколько чужеродных аллелей попадут в генофонд, это, скорее всего, не помешает процессу естественного отбора. Кроме того, в рамках программы DBBB (см. ниже) в период спаривания на пасеках организуют вывод собственных зрелых трутней в избыточном количестве, поэтому свои трутни, скорее всего, легко превзойдут далеко летающих «иностранцев».
3. Отказ от борьбы против клеща V. destructor
На пасеке, работающей по программе DBBB, не предпринимают никаких мер по контролю численности V. destructor на протяжении всего периода действия программы, за исключением одной обработки щавелевой кислотой в первое лето. В то же время контрольную группу семей продолжают лечить от Варроа с целью поддержания численности клеща на приемлемом уровне, чтобы снизить ущерб и исключить давление отбора* под влиянием V. destructor. В нашем случае достаточным методом контроля Варроа мы считаем обработку пчёл щавелевой кислотой два раза в год до запечатывания расплода (Panziera et al., 2017).
* Давление отбора — показатель интенсивности действия естественного отбора с точки зрения изменения генетического состава популяций в ряду поколений. Чем интенсивнее отбор, тем выше скорость эволюционных изменений в популяции. Количественно давление отбора оценивается по величине изменения аллельных частот в популяции за 1 поколение (ru.wikipedia.org, прим. переводчика).
4. Подкормка пчёл
Безусловно, подкормка пчёл не естественна. Из работ Сили (Seeley, 1995, 2017) мы знаем, что в дикой природе (в данном исследовании – Арнотский лес, США) только 23% роев-перваков, успешно нашедших себе новое гнездо, выживают в первую зиму. У семей, которым удалось успешно прижиться и набрать силу, зимняя выживаемость увеличивается до 84% (Seeley, 2017). Основной причиной гибели пчелосемей в дикой природе может быть недостаток корма и голод. Для сравнения, в Германии на пасеках с хорошим уходом ( = хорошая подкормка и лечение пчёл от V. destructor) зиму переживают 95% семей (Genersch et al., 2010). В нашем подходе мы стараемся избегать дополнительного давления отбора, возникающего из-за недостатка корма, путём подкормки молодых семей сахарным сиропом. Семьи также подкармливаются в периоды плохого взятка.
5. Минимальная продолжительность программы – 4 года
Чтобы увидеть первые результаты, необходимо следовать программе не менее 4 лет, и желательно продлить её на более длительный срок. В одном из предыдущих исследований в Нидерландах мы увидели первые положительные результаты, связанные с изменением динамики размножения V. destructor в необработанных от клеща пчелосемьях, через 4 года (Panziera et al., 2017; Kruitwagen et al., 2017). Недавние исследование Авалоса (Avalos et al., 2017) показало, что у медоносной пчелы, для которой характерна сильная полиандрия, эволюция путём прохождения через «бутылочное горлышко»* может быть действительно очень быстрой (всего десять поколений/лет).
* Эффект «бутылочного горлышка» — сокращение генофонда (то есть генетического разнообразия) популяции вследствие прохождения периода, во время которого по различным причинам (в случае с пчёлами – в результате нашествия клеща Варроа) происходит критическое уменьшение её численности, в дальнейшем восстановленное (ru.wikipedia.org, прим. переводчика).
Принципы
Отказ от борьбы против V. destructor: на пасеке, работающей по программе DBBB, любые меры по контролю клеща прекращают после первого лета. В конечном итоге, только адаптация пчёл к новому паразиту является работающим вариантом (включая, возможно, адаптацию клещей, см. Seeley, 2007), всё остальное – тупиковый путь.
Репродуктивная способность: в регионах умеренного климата дикие колонии A. mellifera будут, если позволяют обстоятельства, размножаться весной или летом, отпуская рой-первак и выращивая новых маток в оставшейся семье (некоторые из которых покинут семью с последующими роями).
Мы следуем этому естественному ходу развития пчелосемей в течение сезона: наши семьи имеют возможность размножаться (выводить трутней и строить роевые маточники), когда захотят. Однако, поскольку мы нацелены на выведение из каждой семьи четырёх новых семей (каждую – с молодой маткой из материнской семьи), мы стараемся избегать потери роёв путём создания отводков непосредственно до начала процесса естественного роения. С удалением матки с отводком, в оставшейся колонии синхронизируется процесс оттягивания маточников.
Это означает, что для формирования нового поколения используются только те семьи, которые эффективно участвуют в процессе производства рамок с трутневым расплодом. Таким образом, отбор будет также зависеть от репродуктивных способностей (производства трутней и матки).
Способность к росту: отбор на репродуктивные способности (описанный выше) косвенно влияет на способность семей наращивать силу весной, так как только те семьи, которые хорошо развиваются, входят в роевое состояние. Отбор на способность к росту имеет большое значение и в течение лета: у молодых семей, возникших в начале лета, есть только три месяца, чтобы вырасти до полноценной семьи, способной пережить следующую зиму.
Выживаемость: Наиболее важный фактор выживания семей – это успешное спаривание маток с несколькими здоровыми трутнями. Те семьи, которые производят сильную и здоровую популяцию зимних пчел, будут иметь хорошие шансы пережить зиму и нарастить силу следующей весной.
План действий в течение года
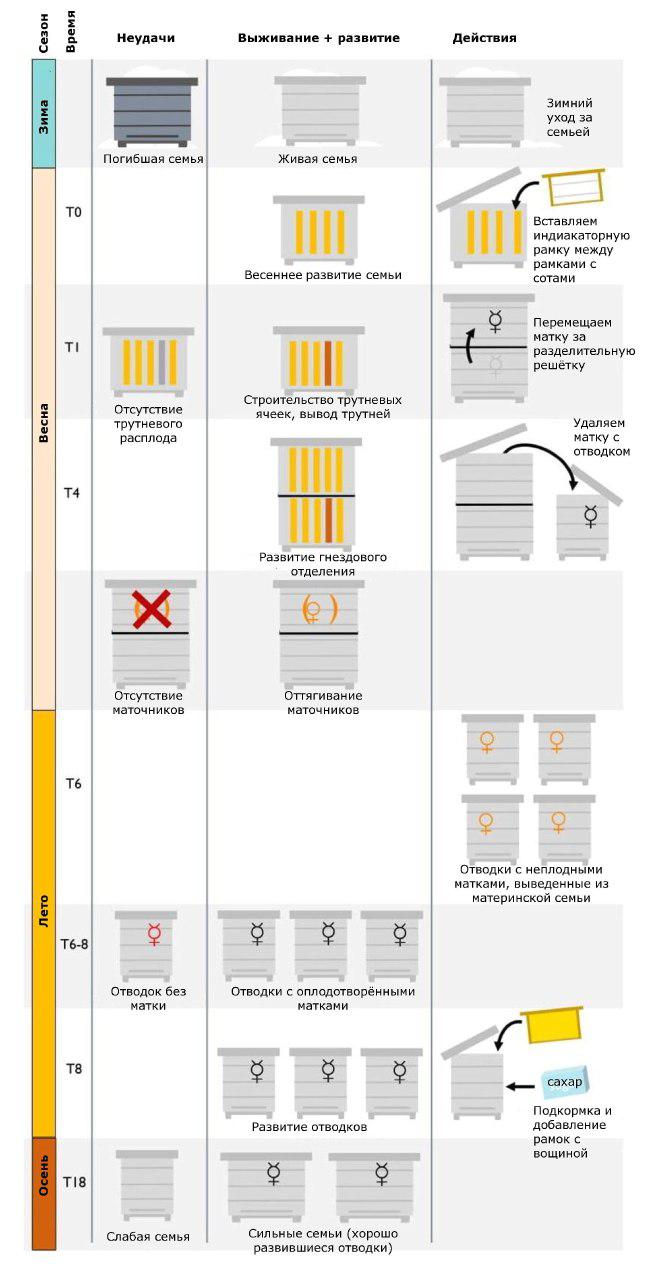
Действия, предпринимаемые пчеловодом в течение года (рис. 1), максимально согласованы с естественным развитием семей. Однако одновременно они должны обеспечивать появление на пасеке достаточного количества новых семей каждый сезон, которые должны компенсировать потери в результате естественного отбора, при этом наиболее важным фактором отбора является способность семей выживать в условиях заражения клещами V. destructor.
[Рисунок 1. Программа Пчелиный Черный Ящик Дарвина по выработке естественной устойчивости к клещу.]
Рисунок 1. Годовой график программы DBBB по неделям, начиная со времени Т0 (= время начала цикла размножения, размещение в ульях рамки-индикатора – рамки с проволокой, но без вощины), с отображением успешных (средний столбец) и неудачных (левый столбец) вариантов развития.
Эти шаги не являются строго обязательными, но они дают природным процессам широкий простор, и природа берёт лидерство в процессе отбора: нежизнеспособные фенотипы погибают после спаривания, до и во время зимы, а также весной; практика пчеловодства насколько возможно учитывает естественное развитие семей в течение сезона; семьи самостоятельно выращивают себе маток.
В контрольной группе семей процесс адаптации пчёл к клещу V. destructor прерывается дважды в год противоклещевой обработкой щавелевой кислотой: летом, через 2 недели после размножения семей отводками, и зимой, когда нет расплода. В зависимости от местных условий и практики могут использоваться и другие известные эффективные методы лечения (обзор методов см. у Rosenkranz et al., 2010).
Перспективы
Программу DBBB можно реализовать легко и быстро, и она может выявить качества пчёл, ответственные за устойчивость.
Семьи европейского подвида A. mellifera благодаря естественному отбору способны выживать в условиях заражения V. destructor, что было продемонстрировано в нескольких случаях (Seeley, 2007; Le Conte et al., 2007; Fries et al., 2006 в обзоре Locke, 2016; Oddie et al., 2017). С тех пор было опубликовано несколько работ, описывающих примеры устойчивости к клещу, приобретённой в условиях культурного пчеловодства (Panziera et al., 2017; Kruitwagen et al., 2017; Kefuss et al., 2016; Oddie et al., 2017; McMullan, 2018).
Во всех примерах критериями отбора были результаты (выживаемость и жизнеспособность пчёл = хорошо развивающиеся семьи) или (медленный) рост популяции клещей, а не отобранные признаки, такие как гигиеническая чувствительность к клещу или активная самоочистка пчёл от клеща. В том числе в исследовании Панциры (Panziera et al., 2017), где использовался именно тот подход, который описан в этой статье.
Популяции медоносных пчёл, полученные в результате такого отбора, по-видимому, приобрели некоторую резистентность и/или толерантность к V. destructor и могут предоставить хорошую возможность раскрыть фенотипические черты, лежащие в основе адаптации к этому новому паразиту на уровне пчёл и семей. Некоторые признаки, эффективные на уровне отдельной пчелы, можно легко отследить на трутнях, поскольку трутни являются гаплоидными (имеющими один набор хромосом). Что еще более важно, такие знания могут помочь нам понять, как вообще развивается резистентность к паразитам у медоносных пчёл.
Используя предложенный подход, мы можем ожидать значимых результатов уже в ближайшее время (4-10 лет), о чём свидетельствуют более ранние исследования (Panziera et al., 2017; Kefuss et al., 2016; Oddie et al., 2017, 2018; McMullan, 2018; Guarna et al., 2017). Если вести также отбор трутней, что является одним из правил данного подхода, то отбор может протекать ещё быстрее, потому что генетически восприимчивые к клещу трутни будут нежизнеспособны и не смогут спариваться, а также потому что сильные семьи выводят больше трутней (Jandricic and Otis, 2003).
Хороший пример быстрого «отрицательного» отбора (исключение неподходящих фенотипов, в данном случае – чрезмерно агрессивных семей) был зарегистрирован в Пуэрто-Рико, где африканизированные пчёлы всего за одно десятилетие стали такими же миролюбивыми, как и европейские (Avalos et al., 2017).
Описанный подход легко могут взять на вооружение непрофессиональные пчеловоды: он прост и не требует специального оборудования. Поэтому его может использовать отдельный пасечник или небольшая группа местных пчеловодов-любителей. Согласно подсчётам (Jandricic and Otis, 2003), данная программа также дешевле, чем любая программа по целевому отбору. Рэнди Оливер (Randy Oliver, 2018), коммерческий пчеловод из Калифорнии, начал сопоставимую программу, включающую предварительный отбор, и завершил её с аналогичными результатами. Голландская коммерческая пчеловодческая компания Inbuzz смогла выполнить такую программу вместе с десятью увлечёнными пчеловодами в течение как минимум 8 лет (Van Stratum, 2016).
Потенциальные ограничения программы DBBB
Желательно, чтобы данный подход использовали в отношении лишь небольшой части местной популяции пчёл: это позволит снизить экономические потери в первые годы. Большинство своих семей пасечники могут продолжать содержать, как и прежде. Однако они могут выращивать новых маток из семей, участвующих в программе отбора, и таким образом постепенно заменить всю популяцию. Преимущество естественного отбора состоит ещё и в том, что адаптированные семьи не потребуют затрат на поддержание здоровья.
Коммерческое матководство в его нынешнем виде не укладывается в предложенный подход, поскольку основой отбора является семья в полном составе. Тем не менее, пчеловоды, использующие наш подход, могут производить дополнительных маток (в большинстве случаев каждая семья выводит больше 4 маток), добавлять их к мини-отводкам на маточник и позволять этим маткам спариваться с остальной популяцией в том же изолированном месте.
Существует вероятность проявления эффекта «бутылочного горлышка», в случае если из первоначальной популяции выживет небольшое количество семей. В любом случае мы не рекомендуем вам полностью изолироваться. Поэтому некоторые чужеродные гены, скорее всего, попадут в популяцию. Если инбридинг все же произойдет, то для сохранения генетического разнообразия потребуется добавить новые семьи.
Существуют фундаментальные биологические различия между восточными (A. cerana) и западными (A. mellifera) медоносными пчелами в отношении клеща V. destructor (см. обзор у Rosenkranz et al., 2010). Например, в семьях пчелы A. cerana клещ успешно размножается исключительно в трутневом расплоде, что нельзя сказать относительно пчёл A. mellifera.
Поэтому очевидно, что новый хозяин клеща A. mellifera вряд ли достигнет точно такого же уровня резистентности/толерантности, как и прежний. Тем не менее, зарегистрированные уровни естественной устойчивости к клещу в европейских подвидах медоносных пчёл (см. выше) в сочетании с очевидной устойчивостью диких и культурных популяций африканизированных и африканских медоносных пчёл (см. обзор у Locke, 2016) убедительно свидетельствуют о том, что фактический уровень устойчивости, достижимый для A. mellifera, по-видимому, достаточен для поддержания стабильных популяций.
Программа DBBB работает на местном, «естественном» масштабе, что поможет избежать распространения неместных болезней и паразитов
Хотя трутни и неплодные матки могут пролетать несколько километров для спаривания, особенно при низкой плотности трутней на территории (Neumann et al., 1999a, b; Jensen et al., 2005), более 75% спариваний происходит с трутнями из околопасечного пространства площадью 4,5 км2 (Jaffe et al., 2009). По оценкам, при плотности размещения 2,4-3,2 пчелосемей на км2 в Германии (Moritz et al. 2007) в пределах досягаемости маток оказываются 11-15 семей, поставляющих трутней. По данным Яффе (Jaffe et al., 2009), среднее медианное количество семей, размещающихся в пределах досягаемости неплодных маток, составляет 10-37 в Европе (10-17 в Германии).
Таким образом, стартуя с 25 семей и имея из-за частичных потерь ~ 15 оставшихся семей, которые следующей весной начнут хорошо развиваться, мы получаем популяцию/плотность семей, очень похожую на ситуацию в природе [хотя в Африке и Средиземноморье численность популяций выше (Jaffe et al. 2009)]. Размер популяций должен быть достаточно большим, чтобы избежать инбридинга, и потенциальный эффект «бутылочного горлышка» будет компенсирован балансирующим отбором* по половому локусу.
* Балансирующий отбор – это такой отбор, результатом которого оказывается не аллельное замещение, а, наоборот, сохранение и повышение разнообразия генов и генотипов в популяции в ходе микроэволюционного процесса. Балансирующий отбор поддерживает в популяции разнообразие аллелей генов, которые дают возможность использовать несколько альтернативных стратегий выживания, обеспечивая поддержание устойчивости всей популяции в условиях действия нескольких агрессивных факторов, влияние которых может быть неравномерным в пространстве и во времени.
Это приведёт к увеличению частоты редких половых аллелей, как это было зарегистрировано у A. cerana после её вторжения в Австралию (Gloag et al., 2016; Ding et al., 2017). Дополнительным преимуществом локальной работы является возможность адаптации к местной среде обитания и сезонности местной кормовой базы (Strange et al., 2007), а также к местным разновидностям болезней (Blacquie`re and Panziera, 2018). Недавний эксперимент в Европе, в котором семьи пчёл разного происхождения территориально меняли местами, показал, что в любом регионе местные семьи всегда работают лучше, чем привезённые, благодаря сильному взаимодействию генотипа и окружающей среды (Büchler et al., 2014; Meixner et al., 2014).
Важно учитывать, что вторжение паразитов и болезней в большинстве случаев происходило в результате перемещения пчёл и маток (Mutinelli, 2011; Owen, 2017). В работе Брози (Brosi et al., 2017) даётся объяснение, как подобная практика снижает иммунитет и повышает уязвимость пчелосемей. Поскольку в нашем подходе сделан акцент на работу на местном уровне, шансы на импорт или экспорт болезней и паразитов в конечном итоге снижаются.
Программа DBBB может увеличить и сохранить функциональное генетическое разнообразие популяций медоносных пчел
Как было обнаружено, на крупных пасеках западная медоносная пчела имеет очень высокое генетическое разнообразие, более высокое, чем в «исходных» эндемичных популяциях Европы (Harpur et al., 2012; Oldroyd, 2012). Это вызвано смешением семей разных европейских подвидов, например, пчёл линии М (А. mellifera Iberica и А. mellifera mellifera) и линии С (А. mellifera carnica и A. mellifera ligustica) (Honey bee Genome Sequencing Consortium 2006).
Хотя столь высокое разнообразие может расцениваться как полезное, на самом деле оно может представлять угрозу для местных акклиматизированных подвидов или популяций. Слишком высокое аллельное разнообразие может замедлить балансирующий отбор по полигенным признакам. И наоборот (хотя одно другое не исключает), спаривание местных акклиматизированных маток с трутнями из неакклиматизированных семей также может увеличить риск гибели семьи.
Поэтому Де Ла Руа (De La Ru´a et al., 2013) настоятельно призывает соблюдать строгие меры, нацеленные на сохранение функционального генетического разнообразия путём сохранения местных подвидов. Эти меры будут эффективны даже в меньшем (более локальном) масштабе, если предложенный нами подход позволит сохранить местные акклиматизированные популяции пчелиных семей. Это в целом увеличит функциональное генетическое разнообразие медоносной пчелы на уровне страны и континента (которое представляет собой сумму местных различий, в отличие от искусственно поддерживаемого разнообразия ограниченным числом селекционеров).
Программа DBBB может повысить приживаемость роёв, тем самым способствуя возвращению медоносных пчёл в дикую природу
Значительная часть популяций западной медоносной пчелы A. mellifera всегда жила в диких условиях (Африка; Jaffe et al., 2009). Пчёлы в дикой природе испытывают сильное давление отбора, причём не только со стороны V. destructor. Но при этом такое существование может также и снизить определенное давление отбора по сравнению с культурными пасеками (Loftus et al., 2016). В случае если дикая популяция достаточно изолирована от культурных пасек, это может способствовать выработке устойчивости (Seeley 2007; Le Conte et al. 2007).
Рои, улетающие с пасеки в дикую природу, часто вскоре погибают (Thompson et al., 2014), вероятно, потому, что не обладают генами, обеспечивающими выживание в отсутствие лечения от клещей. Однако рои из семей, прошедших на пасеке процесс естественного отбора, выживут в диких условиях с большей вероятностью, потому что они приобрели устойчивость к V. destructor. Таким образом, «Пчелиный чёрный ящик Дарвина» вполне может изменить баланс между культурными и дикими колониями, увеличив шансы на выживание и восстановление популяций диких медоносных пчел.
Программа DBBB может способствовать успешной натурализации Varroa destructor
После акклиматизации инвазивного вида усилия по его целенаправленному контролю и смягчению последствий нашествия — именно этот путь был избран в пчеловодстве как предпочтительный — могут стать более дорогостоящими и трудоёмкими, чем принятие как неизбежность того, что новый вид стал «своим» (Epstein, 2017). Между тем, акклиматизация означает, что инвазивный вид стал частью «нашей» природы, и перед ним также встаёт задача адаптации и приспособления к новой нише в качестве натурализованного вида (Blackburn et al., 2011). В отношении акклиматизировавшегося инвазивного вида Varroa destructor мы предлагаем постепенно отойти от мер прямого контроля и передать инициативу в руки естественного отбора. В этой связи мы призываем пчеловодов и ученых следить за нашими исследованиями в рамках программы «Пчелиного чёрного ящика Дарвина», которые начинаются в настоящий момент и в ближайшие годы.
Программа DBBB может оказаться эффективной и в отношении других инвазивных паразитических видов
Поскольку эволюция путём естественного отбора – это механизм, универсальный для всех видов, кажется очевидным, что данный подход в целом можно использовать и для других инвазивных видов (например, для Tropilaelaps spp., de Guzman et al., 2017, а также для Nosema ceranae, Fries, 2010). Во многих случаях инвазивных паразитов контролируют с помощью пестицидов, тем самым мешая приспособлению новых хозяев (см. обзор Dunn and Hatcher, 2015). Нашу программу можно с пользой адаптировать к случаям с другими инвазивными паразитами, таким образом, использование естественного отбора может стать универсальным устойчивым методом, помогающим решать проблемы, связанные с акклиматизированными паразитами и биологическими инвазиями в будущем.
Авторы: Tjeerd Blacquière, Willem Boot, Johan Calis, Arrigo Moro, Peter Neumann & Delphine Panziera
Перевод на русский язык: Елена Арифулина
Похожие статьи:
Пасека:день за днём → Пчёлы исчезнут через 30 лет?
Болезни пчёл → Надо ли вырезать трутневый расплод?
Болезни пчёл → Устойчивость клеща варроа к препаратам
Новости пчеловодства → О тупых американцах, наших дорогах и клеще варроа...
Болезни пчёл → Губит клещей пиво
Болезни пчёл → Меры борьбы с вароатозом пчёл
Болезни пчёл → Могут ли пчёлы бороться с клещём сами?
