
Тепловизионным методом в гнездах зимующих пчелиных семей (ApismelliferaL.) изучали связь между плотностью локализации пчел и распределением тепловых полей. Определена температура тела у пчел, находящихся на периферии скоплений в межсотовых пространствах. Обнаружена сезонная изменчивость реагирования зимующих пчел на колебания внешней температуры.
Известно, что медоносная пчела, не обладая способностью к диапаузе, в состоянии переживать длительную зимовку в условиях холодного и умеренного климата. Зимующие пчелы при пониженной локомоторной активности поддерживают температуру, превышающую порог холодового оцепенения, за счет агрегирования в межсотовых пространствах гнезда и регуляции температуры (1, 2). Сведения о динамике тепловых процессов в гнездах пчелиных семей актуальны для понимания используемых ими механизмов терморегуляции и разработки способов оптимизации зимовки. Имеющееся представление о температурных процессах в гнездах зимующих пчел основано преимущественно на результатах локальных измерений температуры различными типами точечных термометров (термопар, термисторов, термосопротивлений), помещаемых в ульи (2-5). Но эти методы не позволяют проанализировать всю совокупность тепловых процессов в пчелином гнезде, измерить температуру отдельной особи, проследить ее реакцию на изменение внешней температуры и перераспределение внутренних тепловых потоков, определить индивидуальный вклад в регуляцию температурного режима в улье.
Целью настоящей работы было изучение тепловых процессов в гнездах пчелиных семей по интенсивности инфракрасного излучения пчел с акцентом на анализ распределения тепловых потоков и разогрева пчел в разные периоды зимовки в зависимости от колебаний внешней температуры.
Методика. Исследование выполнено на пчелиных семьях по 8,2-20 тыс. рабочих пчел (ApismelliferaL.) в каждой (масса 10 тыс. пчел — примерно 1 кг). Семьи содержались в ульях из поликарбоната под открытым небом. Сотовые рамки в ульях соединялись шарнирно по книжному типу для облегчения доступа в разные участки гнезда. Ульи имели съемные передние и боковые стенки.
Для регистрации инфракрасного излучения пчел использовали тепловизоры ИРТИС-2000 (Россия) и ThermaCam SC3000 («ThermaCam», США), первый — с высоким температурным (0,05 °С), второй — с высоким пространственным (1,1 мрад) разрешением, что позволило с высокой точностью контролировать температуру отдельных особей и распределение тепловых потоков по поверхности скопления пчел. Продолжительность каждого сеанса тепловизионной съемки не превышала 5-6 с; коэффициент пересчета показателей яркостной температуры в реальные значения — 1,01.
Результаты. Осенние похолодания стимулировали агрегирование пчел вокруг зоны максимального разогрева (теплового центра). Исходно это может быть место, в котором завершают развитие пчелы в открытых или запечатанных ячейках. На место локализации пчел оказывало также влияние местоположение летка, поскольку их привлекает поступающий через него воздух и, возможно, свет (с освещенностью летковой зоны в весенне-летний период пчелы связывают начало и завершение полетов). Поэтому с осени наблюдалось смещение межсотовых скоплений к летку (рис. 1).
* Работа выполнена при поддержке РФФИ, проект № 07-04-00305.

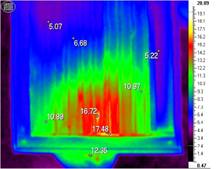
Рис. 1. Вид сверху (слева) и инфракрасная термограмма (справа)надрамочного пространства пчелиной семьи. Масса семьи — 1,8 кг, температура окружающего воздуха — -3 °С.
Анализ тепловизионных термограмм выявил отсутствие внутри гнезда четко выраженного стабильного теплового распределения, которое обычно представляют в виде изотерм. Чередование периодов покоя и активного состояния (теплогенерации) значительно изменяло температурный режим в разных точках скопления пчел (рис. 2). Кроме того, движение особей внутри скопления создавало тепловые потоки, что приводило к локальным изменениям тепловых процессов. При этом с понижением внешней температуры возрастало число активных пчел непосредственно на нижней поверхности межсотового скопления.
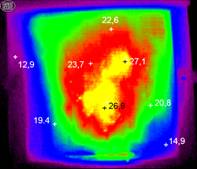
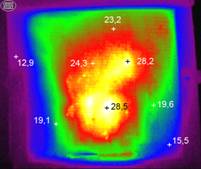
Рис. 2. Термограммы, отражающие изменения тепловых полей в зимнем скоплении пчел при чередовании периода покоя (слева) и термогенерации (справа).
Разогрев разных частей тела у пчел, находящихся на периферии скопления, зависел от места локализации особи и внешней температуры. При ее колебаниях от -1 до -13 °С наибольшую температуру чаще всего регистрировали у грудного отдела, однако среднее значение при этом отличалось от показателей для брюшного и головного отделов на десятые доли градуса (не более 1 °С). Существенно (примерно вдвое) различались средние значения температуры тела у пчел, локализующихся в нижней и верхней частях межсотовых скоплений.
С понижением внешней температуры температура тела у пчел, находящихся на периферии скоплений, изменялась: при внешней температуре -1 °С у 176 особей из верхней части скоплений она равнялась в среднем 22,4±2,4, в нижней — 12,8±3,1 °С. Понижение внешней температуры до -13 °С повлекло усиление разогрева пчел в верхней части скопления в среднем на 1,2, в нижней — на 1,9 °С (Р і 0,99).
Колебания внешней температуры в течение зимовки оказывали наибольшее влияние на разогрев теплового центра. Температура в нем, уменьшаясь от начала к завершению зимовки, варьировала в зависимости от численности пчелиной семьи. Так, при размере семьи около 20 тыс. пчел в начале зимовки колебания температуры в тепловом центре достигали 2,5…2,7, в конце — 1,0…1,6 °С. Коэффициент корреляции между изменением внешней температуры от -17 до +11 °С и разогревом теплового центра уменьшался соответственно с -0,81±0,10 до -0,13±0,04. В семьях, содержавших около 10 тыс. пчел, эта связь была слабее и отмечалась только в начале зимовки (коэффициент корреляции -0,36±0,12).
Распределение пчел в смежных межсотовых пространствах в начале периода зимовки зависело от внешней температуры и числа особей в гнезде. Плотность агрегирования при прочих равных условиях естественной зимовки была максимальной у нижней поверхности скопления и снижалась по мере приближения к тепловому центру (зона максимального разогрева). Относительно невысокой плотностью характеризовалась зона над тепловым центром, что связано с ее обогревом восходящими тепловыми потоками из наиболее теплой части гнезда.
Температурные градиенты от теплового центра к верхней и нижней границе скопления пчел существенно различались по абсолютным значениям и вариабельности: от центра вверх показатели изменялись незначительно (разница в течение зимовки не превышала 12 °С), вниз — постоянно изменяясь, различались на 24…28 °С. При -20…-18 °С в таких же пределах поддерживалась разность между внешней температурой и температурой тела у пчел из нижней части скопления.
Анализ термограммам тепловых потоков пчелиных семей (8,2-19,5 тыс. особей) позволил выявить пределы колебаний внешней температуры, не оказывающие существенного влияния на тепловые потери в скоплении. Было обнаружено, что при температурах от -3,0 до +8,8 °С мощность теплового потока, приходящегося на 1 см2 поверхности межсотового скопления пчел, несущественно зависит от их числа и составляет 2,05-3,21Ѕ10-3 Вт/см2. Это согласуется с результатами изучения воздействия температуры на активность метаболизма, определенную по потреблению кислорода и выделению диоксида углерода (6).
Как оказалось, в отмеченном диапазоне температур семьи, содержащие 14,0±5,5 тыс. пчел, в состоянии поддерживать в тепловом центре необходимую температуру при одинаковой тепловой нагрузке на каждую особь. Увеличение численности пчел сказывалось на тепловых процессах в их скоплениях. В частности, при резких колебаниях внешней температуры наблюдалось увеличение степени температурных флуктуаций в тепловом центре. Такие флуктуации в сочетании с понижением внешней температуры существенно влияют на увеличение теплогенерации каждой пчелой, что сопряжено с дополнительным потреблением корма. Поэтому небольшие семьи при низких температурах погибают не от замерзания пчел (то есть кристаллизации жидких фракций в организме), а от голода, причем нередко гибель происходит даже при наличии в гнезде меда, к которому пчелы не могут приблизиться, израсходовав запасы медовых зобиков и утратив вследствие этого возможность разогреться.
Таким образом, суммарный поток тепла с поверхности скоплений пчел, находящихся в межсотовых пространствах, зависит от ее площади и внешней температуры. У агрегирующих пчел энергетические затраты на обеспечение процессов жизнедеятельности наименьшие при внешней температуре от -3,0 до +8,8 °С. Изменение степени дисперсии (вариабельности) температуры в скоплении от начала к завершению периода зимовки служит показателем адаптивного потенциала пчелиной семьи. Высокая степень дисперсии в начале зимовки соответствует большему числу возможных состояний и вероятности перехода к ним, чем достигается выбор оптимальной стратегии поведения. Снижению разброса флуктуаций внутренней температуры к завершению зимовки соответствует уменьшение возможных вариантов реагирования на изменения внешних условий. Пчелы, характеризующиеся наибольшей активностью, обеспечивают разогрев межсотовых скоплений, что сопряжено с большими затратами энергии. У некоторой части пассивных особей, находящихся на периферии тех зон скопления, которые подвергаются наибольшему охлаждению, затраты энергии минимальны. Эти пчелы, имеющие относительно низкую температуру тела, очевидно, находятся в состоянии неглубокого холодового оцепенения со значительным замедлением обменных процессов.
Е.К. ЕСЬКОВ, В.А. ТОБОЕВ
Источник:http://www.agrobiology.ru
Похожие статьи:
Разведение и содержание пчёл → Размышления о зимовки пчёл
Начинающему пчеловоду → Сахар для зимовки пчёл
Разведение и содержание пчёл → А.М.Бутлеров "О зимовки пчёл"
Методы пчеловождения → Впереди у нас зима
Новости пчеловодства → Еще раз о зимовке
Новости пчеловодства → Зима близка...
Разведение и содержание пчёл → Решение вопроса зимовки
